«Langkah pertama glikolisis
Enzim pertama yang digunakan pada fase kedua glikolisis adalah gliseraldehida 3-fosfat dehidrogenase; dehidrogenase adalah enzim yang mengkatalisis transfer daya reduksi dari molekul pereduksi yang mengoksidasi ke molekul lain yang tereduksi (reaksi redoks).Substrat dari enzim ini adalah NAD (nicotidamide adenine dinucleotide) dan FAD (flavin adenin dinucleotide).
Pada langkah ini dehidrogenase mengkatalisis konversi gliseraldehida 3-fosfat menjadi 1,3-bisfosfogliserat: pada situs katalitik yang sama, gugus aldehida dioksidasi menjadi karboksil dengan konsekuensi reduksi NAD + menjadi NADH dan, selanjutnya, gugus karboksil mampu membentuk ikatan anhidrida dengan ortofosfat. Proses pertama sangat eksergonik (melepaskan energi) sedangkan yang kedua sangat eksergonik (membutuhkan energi); jika tidak ada situs katalitik, reaksi global tidak akan terjadi: reaksi pertama akan terjadi dengan pelepasan energi yang akan terdispersi sebagai panas dan, oleh karena itu, tidak dapat digunakan untuk membentuk ikatan anhidrida.

Setelah pembentukan 1,3-bisfosfogliserat, enzim melanjutkan struktur awalnya dan siap bekerja pada substrat baru.

Kemudian datang kinase fosfogliserat yang memungkinkan transfer fosforil dari 1,3-bifosfogliserat ke molekul ADP; kami telah memperoleh ATP (satu ATP untuk setiap molekul gliseraldehida 3-fosfat, oleh karena itu, dua ATP untuk setiap molekul glukosa awal) yang mengkompensasi pengeluaran energi fase pertama glikolisis.
Anion arsenat (AsO43-) mempengaruhi jalur glikolitik karena dapat menggantikan fosfat dalam reaksi pertama fase kedua glikolisis, memberikan 1-arsenio 3-fosfogliserat yang sangat tidak stabil dan, segera setelah bebas dari katalis. situs, menghidrolisis melepaskan "arsenat kembali ke sirkulasi. Oleh karena itu, arsenat meniru aksi fosfat dan memasuki situs katalitik: dengan adanya arsenat, reaksi yang menghasilkan ATP (dari 1,3-bisfosfogliserat menjadi 3-fosfogliserat) tidak terjadi karena gliseraldehida 3-fosfat diubah langsung menjadi 3-fosfogliserat; tanpa ATP tersedia, sel mati (keracunan asam arsenik).
Pada reaksi ketiga fase oksidatif, 3-fosfogliserat diubah menjadi 2-fosfogliserat oleh aksi fosfogliserat mutase; reaksi melibatkan zat antara 2,3-bifosfogliserat.

Pada langkah selanjutnya, enzim mengintervensi enolase yang mampu mengkatalisis eliminasi molekul air dari kerangka karbon dari 2-fosfogliserat, memperoleh fosfoenol pirivat (PEP);

PEP memiliki potensi tinggi untuk mentransfer fosforil: transfernya, melalui aksi enzim piruvat kinase, sebuah fosforil ke ADP untuk memberikan ATP, pada langkah kelima dari fase kedua, memperoleh piruvat.

2-fosfogliserat dan 3-fosfogliserat memiliki daya transfer fosforil yang rendah oleh karena itu, untuk memperoleh ATP dari molekul-molekul ini, 3-fosfogliserat diubah menjadi 2-fosfogliserat, selama glikolisis, karena diperoleh dari yang terakhir. spesies dengan potensi transfer tinggi.
Sebelum melanjutkan, mari kita buka tanda kurung pada 2,3-bifosfogliserat; yang terakhir hadir di semua sel di mana glikolisis terjadi dalam konsentrasi yang sangat rendah (ini adalah perantara dari reaksi ketiga dari fase kedua glikolisis). Dalam eritrosit, di sisi lain, 2,3-bifosfogliserat memiliki konsentrasi stasioner 4-5 mM (konsentrasi maksimum) karena mereka memiliki warisan enzimatik yang bertugas memproduksinya; dalam eritrosit terjadi penyimpangan dari glikolisis untuk menghasilkan 2,3-bifosfogliserat: 1,3-bisfosfogliserat diubah menjadi 2,3-bifosfogliserat oleh aksi bifosfogliserat mutase (eritrosit) dan 2,3-bifosfogliserat, melalui aksi bifosfogliserat fosfatase (eritrosit) menjadi 3-fosfogliserat. Kemudian, di dalam eritrosit, bagian dari 1,3-bifosfogliserat yang diperoleh dari glikolisis diubah menjadi 2,3-bifosfogliserat yang kemudian kembali ke jalur glikolitik sebagai 3-fosfogliserat; dengan demikian, langkah ketiga dari fase oksidatif glikolisis dari mana ATP diperoleh Jumlah ATP yang hilang adalah harga yang bersedia dibayar oleh eritrosit untuk menjaga konsentrasi 2,3-bifosfogliserat yang dibutuhkan sel-sel ini karena mempengaruhi kemampuan "hemoglobin untuk mengikat" oksigen.
Kita telah melihat bahwa dalam reaksi pertama fase kedua glikolisis, NAD + direduksi menjadi NADH tetapi perlu bahwa, setelah memperoleh piruvat, NADH diubah kembali menjadi NAD +: ini terjadi dengan fermentasi laktat (diperoleh laktat) atau dengan fermentasi alkohol (piruvat dekarboksilase yang mendekarboksilat piruvat dan dehidrogenase yang membentuk etanol ikut berperan); fermentasi tidak melibatkan oksigen (anaerob).
Karena fermentasi laktat, asam laktat, jika tidak dibuang secara memadai, terakumulasi di otot dan, melepaskan H +, menyebabkan kontraksi otot yang tidak disengaja dan, oleh karena itu, kram; otot dalam stres yang kuat juga dapat mencapai pH minimum 6,8 .
Melalui siklus Cori, bagian dari kelelahan otot ditransfer ke hati ketika otot kelebihan beban. Misalkan otot bekerja tanpa suplai oksigen (asumsi yang salah): jika otot bekerja cukup, ATP yang dibutuhkan untuk kontraksi disediakan secara eksklusif oleh glikolisis Jika aktivitas otot meningkat dan ATP tambahan diperlukan, percepat metabolisme aerobik, konversi laktat, yang kemudian dibuang, menjadi glukosa. Pada kenyataannya, otot mengeksploitasi metabolisme aerobik: jika ada ketersediaan oksigen, otot mengeksploitasi, di atas segalanya, ATP yang disediakan oleh metabolisme aerobik dan, ketika tidak ada lagi oksigen yang tersedia, metabolisme anaerobik dipercepat melalui siklus Cori. Siklus ini mengasumsikan bahwa laktat ditransfer dari otot ke hati, di mana, dengan mengeluarkan energi, lebih banyak glukosa diproduksi yang kembali ke otot. Melalui siklus ini, bagian dari ATP yang dikonsumsi di otot disuplai oleh hati yang, melalui proses glukoneogenesis, mampu menghasilkan glukosa yang dapat digunakan oleh otot untuk memperoleh ATP.
Metabolisme glukosa yang dijelaskan sampai sekarang tidak termasuk oksigen tetapi metabolisme glukosa aerobik memungkinkan untuk memperoleh jumlah ATP 17-18 kali lebih tinggi daripada yang diperoleh dengan jalur glikolitik, oleh karena itu, ketika sel memiliki kemungkinan untuk memilih antara aerobik dan ed anaerob, menyukai yang pertama.
Dalam metabolisme aerobik, piruvat memasuki mitokondria di mana ia mengalami transformasi dan akhirnya karbon dioksida dan air diperoleh; dengan cara ini 34 molekul ATP diperoleh untuk setiap molekul glukosa yang terdegradasi.

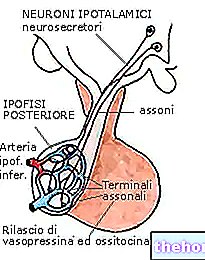
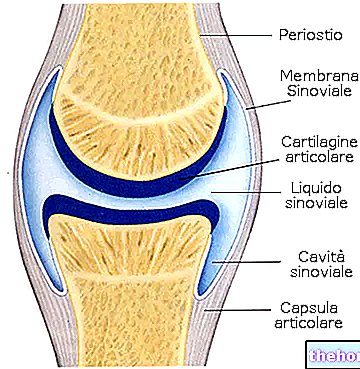




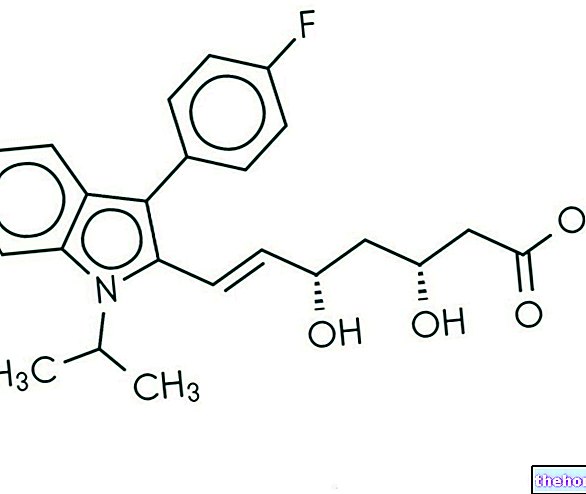









.jpg)