ASOSIASI (ATAU "CONCATENATION" ATAU "LINKAGE")
Sampai sekarang kita telah memperlakukan persilangan dihibrid Mendel (atau polihibrid) dengan asumsi bahwa pasangan alel yang berbeda sebenarnya ditemukan pada pasangan kromosom homolog yang berbeda.Tetapi jumlah pasangan kromosom, meskipun berbeda dari spesies ke spesies, bervariasi dalam batas yang sempit (beberapa spesies mencapai sekitar seratus kromosom), sedangkan jumlah gen dapat dihitung dalam puluhan ribu.
Bahwa karakter-karakter yang dipilih oleh Mendel untuk eksperimen-eksperimennya memisahkan semuanya secara independen (tanpa membingungkan perhitungan dalam distribusi fenotipe dalam F2 persilangan polihibrid) adalah suatu keberuntungan.Jika dua pasang alel ditemukan pada lokus yang berdekatan, hukum bahwa akan mengikuti akan disebut hukum asosiasi.
Mengetahui bahwa sangat banyak karakter memiliki lokus pada satu kromosom dan bahwa mereka adalah pasangan kromosom yang memisahkan secara independen dalam meiosis, dapat dipahami dengan baik seberapa sering terjadi bahwa dua pasang karakter, jika mereka dikaitkan pada kromosom di induk. organisme, tetap berasosiasi, sama juga dalam gamet dan oleh karena itu dalam organisme yang akan membawa materi genetiknya sendiri.
Jadi kita melihat bahwa "asosiasi mewakili" pengecualian, jauh dari jarang, untuk kemerdekaan yang dinyatakan dalam hukum ketiga Mendel.
PERTUKARAN ATAU "CROSS-OVER" DAN REKOMBINASI
Berbicara tentang meiosis, kami telah menunjukkan bahwa ada dua momen pencampuran materi genetik yang berbeda: satu adalah pemisahan kromosom dalam gamet, dan itu adalah yang diamati oleh Mendel.
Momen lain, yang sebenarnya mendahului, adalah momen di mana empat kromatid dari setiap pasangan kromosom homolog saling bertukar sifat identik. Setelah pertukaran ini, dua faktor yang terkait pada kromosom yang sama akan menjadi independen dalam gamet. Probabilitas bahwa pertukaran terjadi sebanding, dengan pendekatan pertama, dengan panjang kromosom, dan dalam kromosom yang lebih panjang bahkan bisa ada lebih dari, pertukaran.
Fenomena ini dapat dideteksi secara sitologi, dengan mengamati jumlah meiosis yang cukup di bawah mikroskop.
Tingkat kecocokan ulang adalah tingkat di mana dua karakter yang terkait dalam generasi induk digabungkan kembali secara berbeda di F2.
Jika kedua lokus tersebut benar-benar bersebelahan, kemungkinan bahwa kiasma akan memisahkan mereka akan praktis nihil. Tingkat rekombinasi akan menjadi: n ° rekombinan. Jika dua lokus berada pada dua kromosom yang berbeda, tingkat rekombinasi akan menjadi 0,5 (probabilitas yang sama, untuk dua karakter yang tergabung dalam generasi P, untuk secara acak menemukan diri mereka bersama di F2). Oleh karena itu, tingkat rekombinasi dapat bervariasi antara 0,0 dan 0,5. Untuk jarak kecil pada kromosom, jarak dan laju rekombinasi berbanding lurus. Untuk jarak yang lebih jauh ada kemungkinan bahwa dua pertukaran terjadi antara dua lokus. Sekarang akan tampak jelas bahwa dua faktor yang dipisahkan oleh dua pertukaran sekali lagi terkait. jelas, pada titik ini, bahwa proporsionalitas antara jarak lokus dan probabilitas rekombinasi hilang.
Lokus yang ditemukan terkait pada kromosom yang sama merupakan 'kelompok asosiasi'. Lokus yang sangat jauh mungkin memiliki probabilitas pemisahan melalui pertukaran sehingga mereka berperilaku sebagai independen, tetapi masing-masing dari mereka akan dikaitkan, dengan tingkat rekombinasi yang lebih rendah, ke lokus perantara.
Ketika tingkat rekombinasi antara banyak pasangan gen dalam kelompok asosiasi diketahui, konstruksi 'peta genetik' dapat dimulai. Mengingat bahwa jarak antara dua gen (a dan b) dinyatakan dengan laju rekombinasi dan bahwa jarak a dari gen ketiga c dapat berupa jumlah atau selisih terhadap jaraknya dari b, adalah mungkin untuk merekonstruksi peta jarak timbal balik, yang akan menjadi peta genetik dalam kelompok asosiasi itu, yaitu kromosom itu.
Kita sekarang harus mempertimbangkan secara umum beberapa konsep yang membatasi manifestasi fenotipik karakter genotip.
Pertama-tama kita akan berbicara tentang konsep penetrasi dan ekspresif, dan kemudian kita akan mencurahkan perhatian khusus pada fenomena regulasi aksi gen.
PENETRASI
Penetrasi gen mewakili kemampuannya untuk memanifestasikan dirinya dalam fenotipe. Penetrasi diukur secara statistik dengan menghitung frekuensi fenotipe yang menunjukkan karakter tersebut dari 100 genotipe yang mengandungnya. Sifat dengan penetrasi 0,7 adalah sifat yang terjadi secara fenotip pada 70% frekuensi genotipnya.
EKSPRESIVITAS
Ekspresivitas adalah penilaian kuantitatif derajat manifestasi fenotipik.
PERATURAN AKSI GEN
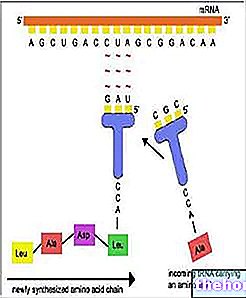
Sel menghasilkan semua enzim dan proteinnya pada kecepatan dan waktu yang sama. Sel Escherichia coli, misalnya, dapat disuplai dengan energi dan atom karbon dari disakarida laktosa karena dapat memecahnya menjadi glukosa dan galaktosa berkat enzim beta-galaktosidase.Pada E.coli normal yang mungkin memiliki laktosa, ada sekitar 3.000 molekul beta-galaktosidase, sama dengan 3% dari protein sel itu; tanpa adanya laktosa hanya akan ada satu molekul beta-galaktosidase per sel bakteri. Galaktosidase akan disintesis dari molekul mRNA baru ketika dapat digunakan. Strain mutan E. coli yang kaya akan enzim diketahui bahkan ketika laktosa tidak ada: mutan ini kurang beruntung dibandingkan dengan sel normal karena mereka dipaksa untuk mengkonsumsi energi dan bahan yang tidak perlu untuk menghasilkan enzim yang akan tetap tanpa substrat. Zat yang menyebabkan peningkatan jumlah enzim, seperti halnya dengan laktosa, akan disebut penginduksi, sedangkan enzim akan dikatakan dapat diinduksi. Zat lain menginduksi, juga dengan cara tertentu, produksi enzim tertentu. Juga pada E. coli, misalnya, mampu membangun semua asam aminonya, yang memiliki karbon dan amonium (NH3), kehadiran asam amino tertentu dalam media kultur (histidin, misalnya) menghalangi produksi semua enzim yang terkait dengannya. biosintesis asam amino itu sendiri: akan dikatakan enzim-enzim ini bahwa mereka dapat ditekan.Dalam sel bakteri, molekul mRNA dihancurkan segera setelah pembentukannya, dan itulah sebabnya mengendalikan produksi mRNA berarti mengendalikan sintesis enzim pada saat yang sama waktu.
OPERA
Untuk menjelaskan bagaimana sel bakteri mampu mengendalikan produksi enzimnya sendiri Jacob dan Monod merumuskan hipotesis operon; operon dibentuk oleh beberapa gen yang secara fungsional terkait dan selaras tanpa diskontinuitas sepanjang bentangan DNA.Operon terdiri dari tiga jenis gen yang berbeda: promotor, tempat pembentukan mRNA dimulai; operator, di mana kontrol dilakukan; satu atau lebih gen struktural, yang mengkode enzim atau protein lain. Dalam sistem beta-galaktosidase, operon mencakup, selain untuk beta-galaktosidase, juga dua gen lain yang mengkode struktural untuk enzim lain yang terlibat dalam metabolisme laktosa.Gen-gen ini berdekatan satu sama lain dan ditranskripsikan satu demi satu sepanjang heliks DNA yang sama menjadi molekul mRNA tunggal.Molekul mRNA yang dihasilkan aktif untuk waktu yang sangat singkat, setelah itu mereka dihancurkan oleh enzim tertentu.
Aktivitas operon pada gilirannya dikendalikan oleh gen lain, regulator, yang mungkin juga jauh dari operon: regulator ini mengkodekan protein, yang disebut represor, yang tampaknya mengikat DNA pada gen operator. antara promotor dan gen struktural sebenarnya menghalangi produksi mRNA.
Represor pada gilirannya dikendalikan, dan kontrol dilakukan melalui zat "sinyal". Dalam kasus enzim yang dapat diinduksi zat ini adalah "induser. Induser" mengikat molekul represor memodifikasi bentuknya sehingga tidak dapat lagi beradaptasi dengan DNA: dalam hal ini, karena tidak ada represor antara promotor dan gen struktural , penekan dapat membentuk molekul mRNA dan dari molekul protein ini. Dengan habisnya suplai penginduksi lagi, regulator akan mendapatkan kembali kendali, yang akan menghentikan produksi mRNA baru, oleh karena itu protein baru. Dalam sistem beta-galaktosidase, penginduksinya adalah laktosa atau zat yang sangat mirip dengan ini. akan bergabung dengan represor untuk menonaktifkannya sehingga memungkinkan biosintesis enzim. Dalam kasus enzim yang dapat ditekan, zat yang bertindak sebagai "sinyal" bertindak sebagai korepresor: penekan hanya aktif jika dikombinasikan dengan korepresor. Dalam sistem histidin, yang melibatkan selusin enzim yang berbeda, asam amino inilah yang dikombinasikan dengan tRNA-nya, korepresor histidin.
INTERAKSI ALOSTERIS
Interaksi alosterik, yang melibatkan inaktivasi enzim dengan mengubah bentuknya, memberikan cara yang berbeda untuk mengatur aktivitas metabolisme sel. Interaksi alosterik memungkinkan kontrol yang lebih akurat daripada sistem induktor-represor operon, tetapi tidak mencapai hasil yang berguna dengan mengecualikan biosintesis zat tertentu dari tahap pertama - produksi mRNA.
SISTEM KONTROL DI EUCARIUS
Ada beberapa fakta yang mengarah pada kepercayaan bahwa sistem regulasi yang mirip dengan operon beroperasi dan unggul di antara tumbuhan dan hewan. Kromosom organisme ini sangat berbeda dari E. coli dan prokariota lainnya. sel sangat berbeda.Mekanisme mitosis sedemikian rupa sehingga setiap sel tumbuhan atau hewan tertentu memiliki semua informasi
genetika hadir dalam telur yang dibuahi. Oleh karena itu, sebagian besar gen dalam sel khusus mana pun akan tetap tidak efisien sepanjang hidup sel. DNA dalam sel-sel ini selalu dikaitkan dengan protein. Jadi ada kemungkinan bahwa represi gen pada eukariota membutuhkan asosiasi ini secara tepat antara DNA dan protein.